Animal heme-dependent peroxidases
| Animal heme-dependent peroxidase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
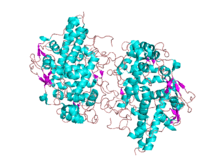 Crystal structure of the human myeloperoxidase-thiocyanate complex.[1] | |||||||||
| Identifiers | |||||||||
| Symbol | An_peroxidase | ||||||||
| Pfam | PF03098 | ||||||||
| InterPro | IPR002007 | ||||||||
| PROSITE | PDOC00394 | ||||||||
| SCOP | 1mhl | ||||||||
| SUPERFAMILY | 1mhl | ||||||||
| OPM superfamily | 37 | ||||||||
| OPM protein | 1q4g | ||||||||
| CDD | cd05396 | ||||||||
| |||||||||
Animal heme-dependent peroxidases is a family of peroxidases.
Peroxidases are found in bacteria, fungi, plants and animals. On the basis of sequence similarity, a number of animal heme peroxidases can be categorized as members of a superfamily: myeloperoxidase (MPO); eosinophil peroxidase (EPO); lactoperoxidase (LPO); thyroid peroxidase (TPO); prostaglandin H synthase (PGHS); and peroxidasin.[2][3][4]
Function
Myeloperoxidase (MPO) plays a major role in the oxygen-dependent microbicidal system of neutrophils. EPO from eosinophilic granulocytes participates in immunological reactions, and potentiates tumor necrosis factor (TNF) production and hydrogen peroxide release by human monocyte-derived macrophages.[5][6] MPO (and possibly EPO) primarily use Cl−ions and H2O2 to form hypochlorous acid (HOCl), which can effectively kill bacteria or parasites. In secreted fluids, LPO catalyses the oxidation of thiocyanate ions (SCN−) by H2O2, producing the weak oxidizing agent hypothiocyanite (OSCN−), which has bacteriostatic activity.[7] TPO uses I− ions and H2O2 to generate iodine, and plays a central role in the biosynthesis of thyroid hormones T3 and T4. Myeloperoxidase (PDB: 1dnu), for example, resides in the human nucleus and lysosome and acts as a defense response to oxidative stress, preventing apoptosis of the cell.[1]
Structure
3D structures of MPO and PGHS have been reported. MPO is a homodimer: each monomer consists of a light (A or B) and a heavy (C or D) chain resulting from post-translational excision of 6 residues from the common precursor. Monomers are linked by a single inter-chain disulfide. Each monomer includes a bound calcium ion.[8] PGHS exists as a symmetric dimer, each monomer of which consists of 3 domains: an N-terminal epidermal growth factor (EGF) like module; a membrane-binding domain; and a large C-terminal catalytic domain containing the cyclooxygenase and the peroxidase active sites. The catalytic domain shows striking structural similarity to MPO. The image at the top of this page is an example of Myeloperoxidase 1dnu derived from X-ray diffraction with resolution 1.85 angstrom.[1]
Active site
The cyclooxygenase active site, which catalyzes the formation of prostaglandin G2 (PGG2) from arachidonic acid, resides at the apex of a long hydrophobic channel, extending from the membrane-binding domain to the center of the molecule. The peroxidase active site, which catalyzes the reduction of PGG2 to PGH2, is located on the other side of the molecule, at the heme binding site.[9] Both MPO and the catalytic domain of PGHS are mainly alpha-helical, 19 helices being identified as topologically and spatially equivalent; PGHS contains 5 additional N-terminal helices that have no equivalent in MPO. In both proteins, three Asn residues in each monomer are glycosylated.
Human proteins containing this domain
The following is a list of human proteins containing this domain:[10]
DUOX1; DUOX2; EPX; LPO; MPO; PTGS1; PTGS2; PXDNL; TPO
References
- 1 2 3 PDB: 1dnu; Blair-Johnson M, Fiedler T, Fenna R (November 2001). "Human myeloperoxidase: structure of a cyanide complex and its interaction with bromide and thiocyanate substrates at 1.9 A resolution". Biochemistry. 40 (46): 13990–7. doi:10.1021/bi0111808. PMID 11705390.
- ↑ Nelson RE, Fessler LI, Takagi Y, Blumberg B, Keene DR, Olson PF, Parker CG, Fessler JH (1994). "Peroxidasin: a novel enzyme-matrix protein of Drosophila development". EMBO J. 13 (15): 3438–3447. PMC 395246
 . PMID 8062820.
. PMID 8062820. - ↑ Poulos TL, Li H (1994). "Structural variation in heme enzymes: a comparative analysis of peroxidase and P450 crystal structures". Structure. 2 (6): 461–464. doi:10.1016/S0969-2126(00)00046-0. PMID 7922023.
- ↑ Kimura S, Ikeda-Saito M (1988). "Human myeloperoxidase and thyroid peroxidase, two enzymes with separate and distinct physiological functions, are evolutionarily related members of the same gene family". Proteins. 3 (2): 113–120. doi:10.1002/prot.340030206. PMID 2840655.
- ↑ Kimura S, Hong YS, Kotani T, Ohtaki S, Kikkawa F (1989). "Structure of the human thyroid peroxidase gene: comparison and relationship to the human myeloperoxidase gene". Biochemistry. 28 (10): 4481–4489. doi:10.1021/bi00436a054. PMID 2548579.
- ↑ Spessotto P, Dri P, Bulla R, Zabucchi G, Patriarca P (1995). "Human eosinophil peroxidase enhances tumor necrosis factor and hydrogen peroxide release by human monocyte-derived macrophages". Eur. J. Immunol. 25 (5): 1366–1373. doi:10.1002/eji.1830250535. PMID 7774640.
- ↑ Wever R, Kast WM, Kasinoedin JH, Boelens R (1982). "The peroxidation of thiocyanate catalysed by myeloperoxidase and lactoperoxidase". Biochim. Biophys. Acta. 709 (2): 212–219. doi:10.1016/0167-4838(82)90463-0. PMID 6295491.
- ↑ Fenna RE, Zeng J (1992). "X-ray crystal structure of canine myeloperoxidase at 3 A resolution". J. Mol. Biol. 226 (1): 185–207. doi:10.1016/0022-2836(92)90133-5. PMID 1320128.
- ↑ Picot D, Loll PJ, Garavito RM (1994). "The X-ray crystal structure of the membrane protein prostaglandin H2 synthase-1". Nature. 367 (6460): 243–249. doi:10.1038/367243a0. PMID 8121489.
- ↑ Zamocky M, Jakopitsch C, Furtmüller PG, Dunand C, Obinger C (August 2008). "The peroxidase-cyclooxygenase superfamily: Reconstructed evolution of critical enzymes of the innate immune system". Proteins. 72 (2): 589–605. doi:10.1002/prot.21950. PMID 18247411.
External links
- Dunand C. "PeroxiBase - The peroxidase database". Swiss Institute of Bioinformatics. Archived from the original on 13 October 2008. Retrieved 2008-11-21.
- C Frank. "MolProbity Ramachandran analysis, 1dnu" (PDF). PDB. Retrieved 2008-11-21.