Biopterin-dependent aromatic amino acid hydroxylase
| Biopterin_H | |||||||||
|---|---|---|---|---|---|---|---|---|---|
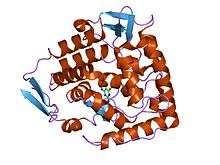 crystal structure of ternary complex of the catalytic domain of human phenylalanine hydroxylase (Fe(II)) complexed with tetrahydrobiopterin and norleucine | |||||||||
| Identifiers | |||||||||
| Symbol | Biopterin_H | ||||||||
| Pfam | PF00351 | ||||||||
| InterPro | IPR019774 | ||||||||
| PROSITE | PDOC00316 | ||||||||
| SCOP | 1toh | ||||||||
| SUPERFAMILY | 1toh | ||||||||
| CDD | cd00361 | ||||||||
| |||||||||
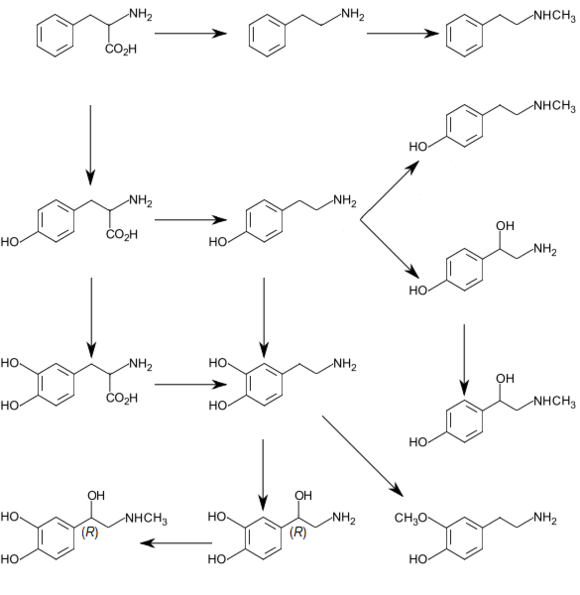
In molecular biology, the biopterin-dependent aromatic amino acid hydroxylases (abbreviated AAAH) constitute a family of aromatic amino acid hydroxylases, including phenylalanine 4-hydroxylase (EC 1.14.16.1), tyrosine 3-hydroxylase (EC 1.14.16.2), and tryptophan 5-hydroxylase (EC 1.14.16.4). These enzymes primarily hydroxylate phenylalanine, tyrosine, and tryptophan, respectively. These enzymes are all rate-limiting catalysts for important metabolic pathways.[1] The proteins are structurally and functionally related, each containing iron, and catalysing ring hydroxylation of aromatic amino acids, using tetrahydrobiopterin (BH4) as a substrate. All are regulated by phosphorylation at serines in their N-termini. It has been suggested that the proteins each contain a conserved C-terminal catalytic (C) domain and an unrelated N-terminal regulatory (R) domain. It is possible that the R domains arose from genes that were recruited from different sources to combine with the common gene for the catalytic core. Thus, by combining with the same C domain, the proteins acquired the unique regulatory properties of the separate R domains.
In humans, phenylalanine hydroxylase deficiency can cause of phenylketonuria, the most common inborn error of amino acid metabolism,[2] Tryptophan hydroxylase catalyzes the rate-limiting step in serotonin biosynthesis: the conversion of tryptophan to 5-hydroxy-L-tryptophan and tyrosine hydroxylase catalyzes the rate limiting step in catecholamine biosynthesis: the conversion of tyrosine to L-DOPA.
References
- ↑ Grenett HE, Ledley FD, Reed LL, Woo SL (August 1987). "Full-length cDNA for rabbit tryptophan hydroxylase: functional domains and evolution of aromatic amino acid hydroxylases". Proc. Natl. Acad. Sci. U.S.A. 84 (16): 5530–4. doi:10.1073/pnas.84.16.5530. PMC 298896
 . PMID 3475690.
. PMID 3475690. - ↑ Erlandsen H, Fusetti F, Martinez A, Hough E, Flatmark T, Stevens RC (December 1997). "Crystal structure of the catalytic domain of human phenylalanine hydroxylase reveals the structural basis for phenylketonuria". Nat. Struct. Biol. 4 (12): 995–1000. doi:10.1038/nsb1297-995. PMID 9406548.
- ↑ Broadley KJ (March 2010). "The vascular effects of trace amines and amphetamines". Pharmacol. Ther. 125 (3): 363–375. doi:10.1016/j.pharmthera.2009.11.005. PMID 19948186.
- ↑ Lindemann L, Hoener MC (May 2005). "A renaissance in trace amines inspired by a novel GPCR family". Trends Pharmacol. Sci. 26 (5): 274–281. doi:10.1016/j.tips.2005.03.007. PMID 15860375.
- ↑ Wang X, Li J, Dong G, Yue J (February 2014). "The endogenous substrates of brain CYP2D". Eur. J. Pharmacol. 724: 211–218. doi:10.1016/j.ejphar.2013.12.025. PMID 24374199.
The highest level of brain CYP2D activity was found in the substantia nigra (Bromek et al., 2010). The in vitro and in vivo studies have shown the contribution of the alternative CYP2D-mediated dopamine synthesis to the concentration of this neurotransmitter although the classic biosynthetic route to dopamine from tyrosine is active. CYP2D6 protein level is approximately 40% lower in the frontal cortex, cerebellum, and hippocampus in PD patients, even when controlling for CYP2D6 genotype (Mann et al., 2012). ... Tyramine levels are especially high in the basal ganglia and limbic system, which are thought to be related to individual behavior and emotion (Yu et al., 2003c). Studies have demonstrated that dopamine is formed from p-tyramine as well as m-tyramine via tyramine 3-hydroxylation or 4-hydroxylation by rat CYP2D2, 2D4, and 2D18 as well as human CYP2D6. ... Both rat CYP2D and human CYP2D6 have a higher affinity for m-tyramine compared with p-tyramine for the generation of dopamine. Rat CYP2D isoforms (2D2/2D4/2D18) are less efficient than human CYP2D6 for the generation of dopamine from p-tyramine. The Km values of the CYP2D isoforms are as follows: CYP2D6 (87–121 μm) ≈ CYP2D2 ≈ CYP2D18 > CYP2D4 (256 μm) for m-tyramine and CYP2D4 (433 μm) > CYP2D2 ≈ CYP2D6 > CYP2D18 (688 μm) for p-tyramine (Bromek et al., 2010; Thompson et al., 2000).
This article incorporates text from the public domain Pfam and InterPro IPR019774