Embryo fossil
Fossil embryos are the preserved remains of organisms that have yet to hatch or be born. Many fossils of the 580 million year old Doushantuo formation have been interpreted as embryos; embryos are also common throughout the Cambrian fossil record.
Preservation
Taphonomic studies indicate that embryos are preserved for longest in reducing, anoxic conditions. These conditions can keep the embryos intact long enough for bacteria to mineralise the cells and permit their preservation.[1] However, phosphatisation was very rapid in the Doushantuo, so it is possible that faster preservation allowed embryos in different regimes to be preserved.
Preservation is mediated by bacterial biofilms. The cells of the embryo break down within hours of their death, under the effect of its own enzymes. Bacteria invade the decaying embryo before its cells can collapse, and biofilms take on and preserve the three-dimensional structure of the cells. The activity of these bacteria promote mineralisation.[2]
Currently, only fossils representing very early stages in embryonic development have been preserved – no embryos have been interpreted as having undergone a large number cleavage stages.[3]
Affinities
Some embryos have been interpreted as colonies of sulfur-reducing bacteria, a claim that cannot be upheld in all cases.[3]
Fossilized embryos of the 635- to 551-million year old Doushantuo Formation of southern China exhibit occasional asynchronous cell division, common in modern embryos, implying that sophisticated mechanisms for differential cell division timing and embryonic cell lineage differentiation evolved before 551 million years ago. However, embryos composed of hundreds to more than ~1000 cells still show no evidence of blastocoel formation or the organization of blastomeres into epithelia - epithelialization should be underway in modern embryos with >100 cells. Features preserved on Doushantuo embryos are compatible with metazoans (animals), but the absence of epithelialization is consistent only with a stem-metazoan affinity.[4] It is not until the Cambrian that embryos with demonstrable eumetazoan features occur; gastrulation and a pentaradial symmetry are expressed in some lower Cambrian fossils.[5]
Some fossil embryos are considered to belong to cnidarians and ecdysozoans, if they even fall into the metazoan crown group. No deuterostome or lophotrochozoan embryos have yet been reported, despite their similar preservation potential; this may be a result of different egg laying behaviour, as ecdysozoans lay eggs in the sediment rather than releasing them into the open – enhancing the chance of them becoming mineralised.[1]
An alternative interpretation of the embryos appears to discount an animal – or indeed embryonic – interpretation altogether. Cell division without enlargement appears to continue beyond what it would in embryos, and without other embryonic traits becoming apparent. Such division is found in a wide range of eukaryotes, including some that are not truly multicellular, and this more conservative interpretation looks to be more parsimonious than embryonic claims.[6]
Some have also been interpreted as algal.[7]
Ediacaran embryos
Helically coiled spheroidal fossil embryos dated to the Ediacaran era were discovered in Doushantuo, in southern China. [8] These fossils are characterised by a three dimensional spiral tunnel that is punctured by a series of[9] pores, or holes. Scientists suggest that these fossils are similar to Megasphaera ornata from aspects such as the similarity in envelope and size, so there is a stated assumption that these fossils represent postblastula embryos of Megasphaera ornata. in addition, There was a remarkable discovery of fossilized eggs and embryos which are believed to be the oldest metazoan fossils known in the fossil record. These fossils show the cleavage stage, and the blastula, but the identity of the adult forms is not yet unknown. Also, there are microfossils considered as one of the largest fossils from the Ediacaran period.[10] These fossils resemble the embryos and larva of metazoans, and clearly show the direct developing lecithotropic forms.
Cambrian embryos
Markuelia embryo fossils from Siberia, and Olivoodes embryo fossils from China are considered as an example of embryo fossils from the earliest Cambrian (541-485.4 million years ago). Markuelia fossils from the Aldan River in southern Yakutia, Siberia show a wormlike animal with chambers. The two ends in one hemisphere are connected to each other. There is a S-shape in the middle part of the fossil. In some specimens there is a rod like structure which is suggested to be lateral nerve branches from a nerve cord that is longitudinal. In addition to earlier and later cleavage stages most Olivooides fossils observed are in a late embryonic stage, and eggs contain an embryo which is looped. The egg fossils show a smooth outer membrane that is covered with nonannulated stellate tissue. Most of these globules have a regular polygonal surface pattern. Some fossils represent early cleavage stages, and others represent the gastrula.[11] Some specimens show a hatched animal which is conical with a star-shape surface, and stellae. They are pertural in the peak. There is an addition of striated tissue which indicates a growth process. These fossils are characterized by a stellate cuticle which covers the surface with regular sharp protrusions. In general, these fossils for (Markuelia, and Olivooides) are direct development, and they represent an early metazoan because of the egg size, which is generally larger in the direct developers compared to eggs of indirect developers. Stellate embryos, cleavage stages, embryos within a fertilization envelope, and early and later post embryonic polyp stages are clearly observed in these fossils. Some Markuelia fossils are currently located in the Swedish Museum of Natural History, while Olivooides fossils are located in the National Geological Museum of China, with some of them deposited at the Chinese Academy of Geological Sciences.
In the Kuanchuaanpu Formation in Shaanxi Province of South China a great number of early Cambrian fossil Cubozoan embryos or "sea wasps" were discovered,[12] (fig 1) marine organisms which are a sister group of the Scyphozoa. "Sea wasps" are characterized by interradial tentacles, four branches, a four-sided box shape, and four well developed complex eyes but no brain. From the entry these species contain four interradial pairs of leaf shaped gonads, and four interradial septa & claustra.[13] Most fossils show the embryo in hemi-spherical shape, and others are ellipsoid. Furthermore, there are two layers to most specimens (exumbrella, and subumbrella), the outer and inner respectively. The gastric groove is represented in between these two layers, and consists of a set of lamellae, and gastric pockets. The main structures that are clear in these fossils are the tentacle which appears as either buds, or primordial, and the mouth cavity. The tentacles that appear in most specimens are between four or five, and they arise from the subumbrellar wall. The oral cavity has lips in some fossils which are around the mouth opening, but in some specimens the oral lips are represented in the perradii. There is a lumen at the end of the oral cavity that represents the esophagus which is an extension of the oral lips. There are straight plates considered as interradial septa which is extended toward the stomach cavity, and there are paired leaf-like gonads lamella.
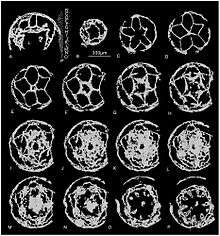
Figure 1: Micro-CT photographs of microscopic cubozoan fossils.
Moreover, in Mackenzie Mountain, Northwest Territories in northwestern Canada metazoan fossils from the early Cambrian have been discovered which appear similar to small circular discs which represent cnidarians, and some creeping bilaterians. A clear presence of guts was noted in bilaterian fossils.
Controversies
These famous fossils of embryos have been discovered in China, in the Ediacaran Doushantuo formation, Datang Quarry, Weng’s, Guizhou Province, southern China. Most of the fossils are characterized by the appearance of palintomic cell cleavage of the nonmetazoa holozoa, a sister group of fungi (fig 2). The study of these fossils has shown that these specimens are related to Tianzhushania (ancestors of Spiralicellula, Megasphaera, and Parapandorina). There are nucleus-like bodies having a globular shape that are clearly shown in all specimens, and their position is central to the cells. In other specimens the area surrounding the nucleus-like bodies is homogeneous, with some finely granular tissue. In the center of these nucleus-like bodies there are spheroidal bodies that represent the nucleoli surrounded by the same tissue. There was an amazing specimen that shows a stage of division because it contains an elongated, dumbbell shape. In addition, there are some fossils that are likely to follow Spiralicellula which is similar to Tianzhushania in terms of the pattern of cell division, and Spiralicellula are characterized by an elongated and vermiform body. This study illustrates how most of these fossils seem to be similar to modern Mesomycetozoeans.

These fossils are still under discussion about their identity, Therese Huldtgren and colleagues said these fossils are incompatible with metazoan multicellular embryos because the developmental pattern resembles that of nonmetazoan holozoans,[6] however, some scientists like Xiao published an article in rebuttal, he said these fossils don’t resemble mesomycetozoeans[14] because the structures that Huldgren said are nuclei are not since the size is too large to be nuclei. Huldtgren responds to this article to say it depends on the number of morphological features whick identify the nuclei such as the position of cells, the volumetric shape between the cells, and the nuclei. She stresses that these fossils don’t represent prokaryotes, multicellular stem or crown metazoans as some believe. In general there are strong controversies and opposition concerning the identity of these fossils, with each scientist trying to prove their theory.
Fossils and Evolution
Records of fossils discovered so far contain a set of examples of people's understanding of the process of biological evolution and help us to answer some questions about evolutionary progression and gives us a unique view into the history of life on earth. Hoatzin chicks; for instance, had claws on their wings, and evidence that the ancestors of birds had clawed digits. Moreover, animals like frogs, rabbits, and lizards have different forelimbs and in spite of the differences, they have the same set of bones with the same arrangement. These bones have the same organisation in the fossil of their common ancestor (Eusthenopteron). Fossils provide us with evidence about processes that happened on our planet, such as the process of transition from one environment to another. For instance scientists in 1988 discovered a special 370 million year-old fossil which is a transitional form from sea to land animals animals (tetrapods). Transitional forms from one species to another exist, such as one reptile group (Cynodonts) which appeared 260 million years ago as reptiles but quickly changed in time to mammaliforms some 245 million years ago. Evolution and the transition process are very clear in the bone structure of the ear, so all fossils that are recorded up to today provide us with sufficient clarity to understand more about evolutionary history.
In general, there is a clear relationship between Evolution (a theory about the history of life), and Paleontology (study of fossils) that provides us with information about how life has changed and developed, and about ourselves as humans.[15] evolution history.
Dinosaurs
Some hadrosaur fossils recovered from the Dinosaur Park Formation may represent embryos.[16]
References
- 1 2 Gostling, N. J.; Thomas, C. W.; Greenwood, J. M.; Dong, X.; Bengtson, S.; Raff, E. C.; Raff, R. A.; Degnan, B. M.; Stampanoni, M.; Donoghue, P. C. J. (2008). "Deciphering the fossil record of early bilaterian embryonic development in light of experimental taphonomy". Evolution & Development. 10 (3): 339–349. doi:10.1111/j.1525-142X.2008.00242.x. PMID 18460095.
- ↑ Raff, E. C.; Schollaert, K. L.; Nelson, D. E.; Donoghue, P. C. J.; Thomas, C. -W.; Turner, F. R.; Stein, B. D.; Dong, X.; Bengtson, S.; Huldtgren, T.; Stampanoni, M.; Chongyu, Y.; Raff, R. A. (2008). "Embryo fossilization is a biological process mediated by microbial biofilms". Proceedings of the National Academy of Sciences. 105 (49): 19360–19365. Bibcode:2008PNAS..10519360R. doi:10.1073/pnas.0810106105.
- 1 2 Gostling, N. J.; Donoghue, P. C. J.; Bengtson, S. (2007). "The earliest fossil embryos begin to mature". Evolution & Development. 9 (3): 206–207. doi:10.1111/j.1525-142X.2007.00152.x.
- ↑ Hagadorn, J. W.; Xiao, S.; Donoghue, P. C. J.; Bengtson, S.; Gostling, N. J.; Pawlowska, M.; Raff, E. C.; Raff, R. A.; Turner, F. R.; Chongyu, Y.; Zhou, C.; Yuan, X.; McFeely, M. B.; Stampanoni, M.; Nealson, K. H. (2006). "Cellular and Subcellular Structure of Neoproterozoic Animal Embryos". Science. 314 (5797): 291–294. Bibcode:2006Sci...314..291H. doi:10.1126/science.1133129. PMID 17038620.
- ↑ Yao, X.; Han, J.; Jiao, G. (2011). "Early Cambrian epibolic gastrulation: A perspective from the Kuanchuanpu Member, Dengying Formation, Ningqiang, Shaanxi, South China". Gondwana Research. 20: 844–851. doi:10.1016/j.gr.2011.04.003.
- 1 2 Huldtgren, T.; Cunningham, J. A.; Yin, C.; Stampanoni, M.; Marone, F.; Donoghue, P. C. J.; Bengtson, S. (2011). "Fossilized Nuclei and Germination Structures Identify Ediacaran "Animal Embryos" as Encysting Protists". Science. 334 (6063): 1696–1699. doi:10.1126/science.1209537. PMID 22194575.
- ↑ Zhang, X. G.; Pratt, B. R. (2014). "Possible Algal Origin and Life Cycle of Ediacaran Doushantuo Microfossils with Dextral Spiral Structure". Journal of Paleontology. 88: 92–98. doi:10.1666/13-014.
- ↑ http://www.ucmp.berkeley.edu/vendian/ediacaran.php
- ↑ Xiao, S. ".Rare helical spheroidal fossils from the Doushantuo Lagerstatte: Ediacaran animal embryos come of age". Geology. 35 (2): 115–118. doi:10.1130/g23277a.1.
- ↑ Raff, E.C. Villinski, J.T. Turner, F.R. Donoghue, P.C.J. and Raff, R.A. 2005. Experimental taphonomy shows the feasibility of fossil embryos. PNAS Journal. 103 (15):5846–5851.
- ↑ Bengtson, S. (1976). "Fossilized Metazoan Embryos from the Earliest Cambrian". Blood. 227 (5332): 91–8. PMID 1126.
- ↑ http://www.ucmp.berkeley.edu/cnidaria/cubozoa.html
- ↑ Han, J (2013). "Early Cambrian Pentamerous Cubozoan Embryos from South China". PLoS ONE. 8: e70741. doi:10.1371/journal.pone.0070741. PMC 3741300
 . PMID 23950993.
. PMID 23950993. - ↑ Xiao, S (2012). "Comment on "Fossilized Nuclei and Germination Structures Identify Ediacaran 'Animal Embryos' as encysting protist"". Science. 335: 1169; author reply 1169. doi:10.1126/science.1218814. PMID 22403373.
- ↑ http://evolution.berkeley.edu/evolibrary/article/evo_01
- ↑ Tanke, D.H.; Brett-Surman, M.K (2001). "Introduction". In D.H. Tanke; K. Carpenter. Evidence of Hatchling and Nestling-Size Hadrosaurs (Reptilia:Ornithischia) from Dinosaur Provincial Park (Dinosaur Park Formation: Campanian), Alberta, Canada. pp. 206-218. Mesozoic Vertebrate Life—New Research Inspired by the Paleontology of Philip J. Currie. Bloomington: Indiana University Press. pp. xviii + 577 pp.
Further reading
- Summary of perspectives up to 2012 (subscription required): Butterfield, N. J. (2011). "Terminal Developments in Ediacaran Embryology". Science. 334 (6063): 1655–1656. Bibcode:2011Sci...334.1655B. doi:10.1126/science.1216125.