Melon (cetacean)


The melon is a mass of adipose tissue found in the forehead of all toothed whales.[1][2] It focuses and modulates the animal's vocalizations and acts as a sound lens. It is thus a key organ involved in communication and echolocation.
Description
The melon is structurally part of the nasal apparatus and comprises most of the mass tissue between the blowhole and the tip of the snout. The function of the melon is not completely understood, but scientists believe it is a bioacoustic component, providing a means of focusing sounds used in echolocation as well as creating a similarity between characteristics of its tissue and the surrounding water so that acoustic energy can flow out of the head and into the environment with the least loss of energy. In the past, some scientists believed that the melon had functions in deep diving and buoyancy, but these ideas have been discounted over the last 40 years and are no longer considered valid by cetologists.[3]
The varying composition of the melon creates a sound velocity gradient that refracts sound directionally. Sounds also bounce off the skull and air sacs that surround the melon.[4]
Melon size is unrelated to maximum dive depth in toothed whales. The particular characteristics of the melon probably have more to do with odontocete phylogeny, the taxonomic relationships over evolutionary time. In some species, melons are more specialized than in others. The sperm whale has the largest nose of any animal in the world. The bulk of that nose is composed of two large, fatty structures, the spermaceti organ and the "junk". The junk is structurally the same as the melon (homologous to it). The melon is not homologous to the spermaceti organ.[1][5]
Composition
The melon is a mixture of triglycerides and wax esters. The exact composition varies throughout the melon. Typically, the inner core of the melon has a higher wax content than the outer parts and conducts sound more slowly. This gradient refracts sound and focuses it like a lens.
The lipids in the melon cannot be digested by the animal as they are metabolically toxic. A starving dolphin will have a robust melon even if the rest of its body is emaciated.[6] The lipids in the melon tend to be of lower molecular weight and more saturated than the blubber.
The melons of Delphinidae (dolphins) and Physeteroidea (sperm whales) have a significant amount of wax ester, whereas those of Phocoenidae (porpoises) and Monodontidae (narwhals and beluga whales) contain little or no wax.[7] The speed of sound in the melon is lowest in the Delphinidae, Phocoenidae and Monodontidae, intermediate in the Ziphiidae, and highest in the Physeteridae and Platanistidae.[8]
Pilot whales
The melon of the pilot whale is a mixture of wax esters and triglycerides. The inner core of the melon is about 33% wax esters, while the outer layer is about 5% wax esters. Most of the fats are saturated.[9]
Pygmy sperm whale
In the pygmy sperm whale (Kogia breviceps), the melon consists of an outer layer and an inner core. The inner core has a generally larger proportion of wax esters than the outer layer.[10]
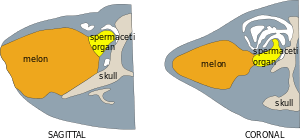
Behind the melon is a cornucopia-shaped organ that many scientists refer to as the "spermaceti organ". This organ is different in form and composition from the spermaceti organ of the sperm whale.
| Outer melon | Inner melon | Spermaceti organ | |
|---|---|---|---|
| Lipid content (weight) | 15-91% | 74-94% | 92-96% |
| Lipid composition | |||
| wax esters | 8-46% | 40-90% | 84-99% |
| triglycerides | 54-92% | 10-69% | 1-16% |
| Average carbon number | |||
| wax esters | 32-35 | 29-32 | 28-29 |
| triglycerides | 47-51 | 41-46 | 45 |
Sperm whale
The analogous structure in the sperm whale is traditionally called "the junk" because whalers dismissed it as a worthless source of sperm oil. It contains compartments of spermaceti separated by walls of cartilage.
Beluga whale
The melon of the beluga whale is also unique in that the whale can change the melon's shape at will.[11] These changes in shape probably have the effect of changing the size, shape, direction, and frequency composition of the echolocation beam.
References
- 1 2 Cranford, Ted W.; Amundin, Mats; Norris, Kenneth S. (June 1996). "Functional morphology and homology in the odontocete nasal complex: Implications for sound generation". Journal of Morphology. 228 (3): 223–285. doi:10.1002/(SICI)1097-4687(199606)228:3<223::AID-JMOR1>3.0.CO;2-3. PMID 8622183.
- ↑ Harper, C.J.; McLellan, W.A.; Rommel, S.A.; Gay, D.M.; Dillaman, R.M.; Pabst, D.A. (July 2008). "Morphology of the melon and its tendinous connections to the facial muscles in bottlenose dolphins (Tursiops truncatus)". Journal of Morphology. 269 (7): 820–839. doi:10.1002/jmor.10628.
- ↑ Cranford, Ted W. (October 1999). "The sperm whale's nose: Sexual selection on a grand scale?". Marine Mammal Science. 15 (4): 1133–1157. doi:10.1111/j.1748-7692.1999.tb00882.x.
- ↑ Marine Mammal Biology: An Evolutionary Approach. pg 153
- ↑ Cranford, T. W., et al. (2008). "Anatomic Geometry of Sound Transmission and Reception in Cuvier's Beaked Whale (Ziphius cavirostris)." Anat Rec 291(4): 353-378.
- ↑ Ted W. Cranford, Mats Amundin, Kenneth S. Norris (1996). "Functional Morphology and Homology in the Odontocete Nasal Complex: Implications for Sound Generation". Journal of Morphology 228:223-285 (1996).
- ↑ Carter Litchfield, R. G. Ackman, J. C. Sipos, C. A. Eaton (1971). "Isovaleroyl triglycerides from the blubber and melon oils of the beluga whale (Delphinapterus leucas)". Lipids, Volume 6, Issue 9, pp 674-681
- ↑ Carter Litchfield, Anne J. Greenberg, David K. Caldwell, Melba C. Caldwell, J.C. Sipos, R.G. Ackman (1975). "Comparative lipid patterns in acoustical and nonacoustical fatty tissues of dolphins, porpoises and toothed whales". Comparative Biochemistry and Physiology Part B: Comparative Biochemistry, Volume 50, Issue 4, 15 April 1975, Pages 591–597
- ↑ Yuri Wedmid, Carter Lichtfield, R. G. Ackman, J. C. Sipos, C. A. Eaton, and E. D. Mitchell. "Heterogeneity of lipid composition within the cephalic melon tissue of the pilot whale (Globicephala Melaena)". Biochimica et Biophysics Acta, 326 (1973) 439-47
- 1 2 R., Karol; C., Litchfield; D., Caldwell; M., Caldwell (1978). "Compositional topography of melon and spermaceti organ lipids in the pygmy sperm whale Kogia breviceps: Implications for echolocation". Marine Biology, Volume 47 (2)
- ↑ "Beluga Whale wiggling its Melon at Vancouver Aquarium". YouTube.
