C10orf118
| CCDC186 | ||||||
|---|---|---|---|---|---|---|
| Identifiers | ||||||
| Aliases | CCDC186, C10orf118, coiled-coil domain containing 186 | |||||
| External IDs | MGI: 2445022 HomoloGene: 9963 GeneCards: CCDC186 | |||||
| RNA expression pattern | ||||||
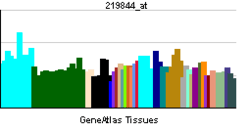 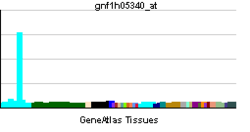 | ||||||
| More reference expression data | ||||||
| Orthologs | ||||||
| Species | Human | Mouse | ||||
| Entrez | ||||||
| Ensembl | ||||||
| UniProt | ||||||
| RefSeq (mRNA) | ||||||
| RefSeq (protein) | ||||||
| Location (UCSC) | Chr 10: 114.12 – 114.17 Mb | Chr 19: 56.79 – 56.82 Mb | ||||
| PubMed search | [1] | [2] | ||||
| Wikidata | ||||||
| View/Edit Human | View/Edit Mouse |

C10orf118 is a protein that in humans is encoded by the C10orf118 gene [4] The C10orf118 (CCDC186) gene is also known as the CTCL-tumor associated antigen with accession number NM_018017.[5]
Gene
Location
C10orf118 has the chromosome location of 10q25.3 and is 53,750 bases in size oriented on the minus strand. PSORTII Protein k-NN Prediction indicated that C10orf118 is 65.2% of the time nuclear, 17.4% cytosolic, 8.7% mitochondrial, 4.3% vesicles of secretory system, and 4.3% endoplasmic reticulum.[6]
Expression
Analysis of gene expression in humans and other species indicates C10orf118 is ubiquitously expressed in all tissue types at varying developmental stages. An EST profile from NCBI displayed the greatest expression in bone marrow, kidneys, and the prostate cell lines. Breakdown by health state indicates high expression of C10orf118 in bladder carcinoma and prostate cancer.[9]
Protein
General Properties
The protein of CCDC186 (NP_060487) is 898 amino acids in length. The predicted molecular weight is 103.7kdal and the isoelectric point is predicted to be 5.92.[10]
Composition
A serine rich region is observed in amino acids 710-747. A compositional analysis revealed that C10orf118 is Proline (1.1%) poor and Glutamic acid (14.1%) and Lysine (12.0%) rich.[10]
Interactions
C10orf118 protein was found to interact with proteins PLEKAH5, Ezra, GAMMAHV.ORF23, and SMAD3.[11][12]
Homology
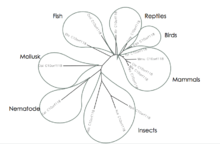
Orthologous sequences of C10orf118 were not found to be in bacteria, archea, protist, or plants. C10orf118 has no human paralogs. The date of divergence for the orthologous sequences highly correlates with the sequences similarity in that the percent identity decreases as you go back in time. Closely related orthologs include mammals and birds and moderately related orthologs include other vertebrates such as fish, reptiles, and amphibians. Distantly related orthologous sequences are primarily observed in invertebrates.[10][14]
| Sequence Number | Genus and Species | Common Name | Date of divergence (MYA) | Accession Number | Sequence length | Seuquence Identity | Sequence Similarity | E-value |
| 1 | Homo sapiens | C10orf118 | 0 | NM_018017 | 898 aa | 100% | 100% | 0% |
| 2 | Nomascus leucogenys | Northern White-cheeked Gibbon | 19.9 | XP_003255542 | 898 aa | 98% | 98% | 0% |
| 3 | Chinchilla lanigera | long tailed chinchilla | 90.9 | XP_013371303 | 906 aa | 90% | 94% | 0% |
| 4 | Mus musculus | house mouse | 90.9 | NP_739563 | 917 aa | 84% | 91% | 0% |
| 5 | Tursiops truncatus | bottlense dolphin | 97.5 | XP_004318425 | 599 aa | 60% | 68% | 0% |
| 6 | Loxodonta africana | African Bush Elephant | 105 | XP_010596062 | 805 aa | 90% | 91% | 0% |
| 7 | Egretta garzetta | little egret | 320.5 | XP_009634185 | 914 aa | 75% | 83% | 0% |
| 9 | Chelonia mydas | geen sea turtle | 320.5 | XP_007056391 | 920 aa | 76% | 83% | 0% |
| 10 | Xenopus tropicalis | western clawed frog | 355.7 | XP_004919435 | 878 aa | 71% | 83% | 0% |
| 11 | Astyanax mexicanus | Mexican tetra | 429.6 | XP_007259323 | 987 aa | 71% | 82% | 5.00E-09 |
| 12 | Oryzias latipes | Japanese rice fish | 429.6 | XP_011486278 | 991 aa | 58% | 71% | 0% |
| 13 | Callorhinchus milii | Australian ghostshark | 482.9 | XP_007903851 | 931 aa | 65% | 79% | 0% |
| 14 | Strongylocentrotus purpuratus | Sea Urchin | 747.8 | XP_011675415 | 884 aa | 39% | 58% | 7.00E-84 |
| 15 | Octopus bimaculoides | California Two-spot Octupus | 847 | XP_014782587 | 1221 aa | 42% | 60% | 3.00E-127 |
| 16 | Aplysiomorpha | sea hare | 847 | Not found in BLAST | 1384 aa | 42% | 63% | 3.00E-145 |
| 17 | Orussus abietinus | wasp | 847 | XP_012288999 | 1057 aa | 36% | 55% | 5.00E-100 |
| 18 | Atta cephalotes | leafcutter ant | 847 | XP_012054311 | 1154 aa | 35% | 55% | 9.00E-93 |
| 19 | Helobdella robusta | leech | 847 | ESO07814 | 997 aa | 29% | 47% | 1.00E-50 |
| 20 | Caenorhabditis elegans | nematode | 847 | NP_871700 | 743 aa | 26% | 34% | 1.00E-25 |
| 21 | Trichoplax adhaerens | NA | 936 | EDV25816 | 976 aa | 26% | 46% | 2.00E-11 |
Motifs
| Motif Information[15] | Position(s) |
| N-glycosylation site | 33-36, 40-43, 109-112, 867-870 |
| cAMP- and cGMP-dependent protein kinase phosphorylation site. | 238-241, 583-586, 794-797 |
| Casein kinase II phosphorylation site. | 2-5, 10-13, 31-34, 36-39, 126-129, 130-133, 135-138, 139-142, 157-160, 228-231, 241-244, 256-259, 405-408, 440-443, 459-462, 490-493, 515-518, 611-614, 617-620, 625-628, 671-674, 690-693, 734-737, 869-872, 879-882 |
| Leucine Zipper Pattern | 588-609, 870-891 |
| N-myristoylation site | 32-37, 75-80, 86-91, 183-188, 721-726, 728-733, 745-750, 810-815, 828-833 |
| Protein kinase C phosphorylation site | 12-14, 35-37, 49-51, 146-148, 228-230, 243-245, 250-252, 278-280, 347-349, 371-373, 405-407, 417-419, 490-492, 500-502, 581-583, 592-594, 596-598, 682-684, 690-692, 825-827, 869-871, 895-897 |
| Tyrosine kinase phosphorylation site | 134-141, 514-522 |
| Cullen Family Profile | 443-552 |
| K-Box Domain Profile | 526-619 |
| Nebulin Repeat Profile | 521-531 |
| Serine Rich Region | 710-747 |
| Rad50 zinc-hook domain profile | 460-562 |
| ATP synthase B/B' CF(0) | 405-433 |
| Alpha-2-macroglobulin RAP, C-terminal domain | 524-705 |
| Tropomyosin | 334-577 |
Post Translational Modification
C10orf118 is predicted to undergo multiple posttranslational modifications including predicted O-beta-GlcNAc attachment, phosphorylation, a nuclear export signal, glycation of lysines, GlcNAc O-glycosylation, N-glycosylation, and NetCorona sites.[16]
Clinical Significance
Prior research indicates that the open reading frame of C10orf118 is linked to cutaneous T-cell lymphoma by a tumor antigen L14-2.[17] The protein C10orf118 is also found at higher than normal levels in the breast cancer cell line BC 8701.[18]
References
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- ↑ http://ibs.biocuckoo.org/online.php
- ↑ Oduru S, Campbell JL, Karri S, Hendry WJ, Khan SA, Williams SC (June 2003). "Gene discovery in the hamster: a comparative genomics approach for gene annotation by sequencing of hamster testis cDNAs". BMC Genomics. 4 (1): 22. doi:10.1186/1471-2164-4-22. PMC 161800
 . PMID 12783626.
. PMID 12783626. - ↑ "Entrez Gene: C10orf118 chromosome 10 open reading frame 118".
- ↑ "PSORT II Prediction". psort.hgc.jp. Retrieved 2016-04-24.
- ↑ "GeneCards".
- ↑ geo. "Home - GEO - NCBI". www.ncbi.nlm.nih.gov. Retrieved 2016-05-08.
- ↑ "Home - UniGene - NCBI". www.ncbi.nlm.nih.gov. Retrieved 2016-04-24.
- 1 2 3 "SDSC Biology Workbench". seqtool.sdsc.edu. Retrieved 2016-04-24.
- ↑ "* in Literature citations". www.uniprot.org. Retrieved 2016-04-24.
- ↑ http://www.uniprot.org/uniprot/Q7Z3E2
- ↑ "Biology WorkBench 3.2".
- ↑ "BLAST: Basic Local Alignment Search Tool". blast.ncbi.nlm.nih.gov. Retrieved 2016-04-24.
- ↑ "Motif Scan". myhits.isb-sib.ch. Retrieved 2016-05-08.
- ↑ "Welcome to CBS". www.cbs.dtu.dk. Retrieved 2016-04-24.
- ↑ de la Peña M, García-Robles I (September 2010). "Intronic hammerhead ribozymes are ultraconserved in the human genome". EMBO Reports. 11 (9): 711–6. doi:10.1038/embor.2010.100. PMC 2933863. PMID 20651741.
- ↑ D’Angelo, M. L. (2013). "Identification of the "Uncharacterized Protein C10orf118" in Breast Cancer Cells and Its Role on the Hyaluronan Metabolism". Matrix Pathobiology, Signaling and Molecular Targets.
- ↑ "Phyre 2 Results for C10orf118_Protein_Structure_". www.sbg.bio.ic.ac.uk. Retrieved 2016-05-08.
Further reading
- Hartley JL, Temple GF, Brasch MA (November 2000). "DNA cloning using in vitro site-specific recombination". Genome Research. 10 (11): 1788–95. doi:10.1101/gr.143000. PMC 310948. PMID 11076863.
- Hartmann TB, Thiel D, Dummer R, Schadendorf D, Eichmüller S (February 2004). "SEREX identification of new tumour-associated antigens in cutaneous T-cell lymphoma". The British Journal of Dermatology. 150 (2): 252–8. doi:10.1111/j.1365-2133.2004.05651.x. PMID 14996095.
- Wiemann S, Arlt D, Huber W, Wellenreuther R, Schleeger S, Mehrle A, Bechtel S, Sauermann M, Korf U, Pepperkok R, Sültmann H, Poustka A (October 2004). "From ORFeome to biology: a functional genomics pipeline". Genome Research. 14 (10B): 2136–44. doi:10.1101/gr.2576704. PMC 528930. PMID 15489336.
- Mehrle A, Rosenfelder H, Schupp I, del Val C, Arlt D, Hahne F, Bechtel S, Simpson J, Hofmann O, Hide W, Glatting KH, Huber W, Pepperkok R, Poustka A, Wiemann S (January 2006). "The LIFEdb database in 2006". Nucleic Acids Research. 34 (Database issue): D415–8. doi:10.1093/nar/gkj139. PMC 1347501. PMID 16381901.
