Glutathione synthetase
| Glutathione synthetase | |
|---|---|
 | |
| Identifiers | |
| Symbol | GSS |
| Entrez | 2937 |
| HUGO | 4624 |
| OMIM | 601002 |
| RefSeq | NM_000178 |
| UniProt | P48637 |
| Other data | |
| EC number | 6.3.2.3 |
| Locus | Chr. 20 q11.2 |
| Eukaryotic glutathione synthase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Human glutathione synthetase | |||||||||
| Identifiers | |||||||||
| Symbol | GSH_synthase | ||||||||
| Pfam | PF03199 | ||||||||
| Pfam clan | CL0483 | ||||||||
| InterPro | IPR004887 | ||||||||
| SCOP | 2hgs | ||||||||
| SUPERFAMILY | 2hgs | ||||||||
| |||||||||
| Eukaryotic glutathione synthase, ATP binding domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
|
Human glutathione synthetase | |||||||||
| Identifiers | |||||||||
| Symbol | GSH_synth_ATP | ||||||||
| Pfam | PF03917 | ||||||||
| InterPro | IPR005615 | ||||||||
| SCOP | 1m0t | ||||||||
| SUPERFAMILY | 1m0t | ||||||||
| |||||||||
| Prokaryotic glutathione synthetase, N-terminal domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Structure of escherichia coli glutathione synthetase at ph 7.5 | |||||||||
| Identifiers | |||||||||
| Symbol | GSH-S_N | ||||||||
| Pfam | PF02951 | ||||||||
| InterPro | IPR004215 | ||||||||
| SCOP | 1glv | ||||||||
| SUPERFAMILY | 1glv | ||||||||
| |||||||||
| Prokaryotic glutathione synthetase, ATP-grasp domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
|
Structure of escherichia coli glutathione synthetase at ph 7.5 | |||||||||
| Identifiers | |||||||||
| Symbol | GSH-S_ATP | ||||||||
| Pfam | PF02955 | ||||||||
| Pfam clan | CL0179 | ||||||||
| InterPro | IPR004218 | ||||||||
| SCOP | 1glv | ||||||||
| SUPERFAMILY | 1glv | ||||||||
| |||||||||
Glutathione synthetase (GSS) (EC 6.3.2.3) is the second enzyme in the glutathione (GSH) biosynthesis pathway. It catalyses the condensation of gamma-glutamylcysteine and glycine, to form glutathione.[2] Glutathione synthetase is also a potent antioxidant. It is found in a large number of species including bacteria, yeast, mammals, and plants.[3]
In humans, defects in GSS are inherited in an autosomal recessive way and are the cause of severe metabolic acidosis, 5-oxoprolinuria, increased rate of haemolysis, and defective function of the central nervous system.[4] Deficiencies in GSS can cause a spectrum of deleterious symptoms in plants and human beings alike.[5]
In eukaryotes, this is a homodimeric enzyme. The substrate-binding domain has a 3-layer alpha/beta/alpha structure.[6] This enzyme utilizes and stabilizes an acylphosphate intermediate to later perform a favorable nucleophilic attack of glycine.
Structure
Human and yeast glutathione synthetases are homodimers, meaning they are composed of two identical subunits of itself non-covalently bound to each other. On the other hand, E. coli glutathione synthetase is a homotetramer.[3] Nevertheless, they are part of the ATP-grasp superfamily, which consists of 21 enzymes that contain an ATP-grasp fold.[7] Each subunit interacts with each other through alpha helix and beta sheet hydrogen bonding interactions and contains two domains. One domain facilitates the ATP-grasp mechanism[8] and the other is the catalytic active site for γ-glutamylcysteine. The ATP-grasp fold is conserved within the ATP-grasp superfamily and is characterized by two alpha helices and beta sheets that hold onto the ATP molecule between them.[9] The domain containing the active site exhibits interesting properties of specificity. In contrast to γ-glutamylcysteine synthetase, glutathione synthetase accepts a large variety of glutamyl-modified analogs of γ-glutamylcysteine, but is much more specific for cysteine-modified analogs of γ-glutamylcysteine.[10] Crystalline structures have shown glutathione synthetase bound to GSH, ADP, two magnesium ions, and a sulfate ion.[11] Two magnesium ions function to stabilize the acylphosphate intermediate, facilitate binding of ATP, and activate removal of phosphate group from ATP. Sulfate ion serves as a replacement for inorganic phosphate once the acylphosphate intermediate is formed inside the active site.[6]
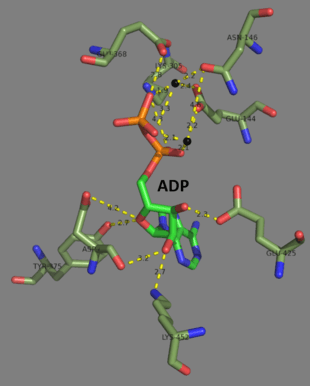
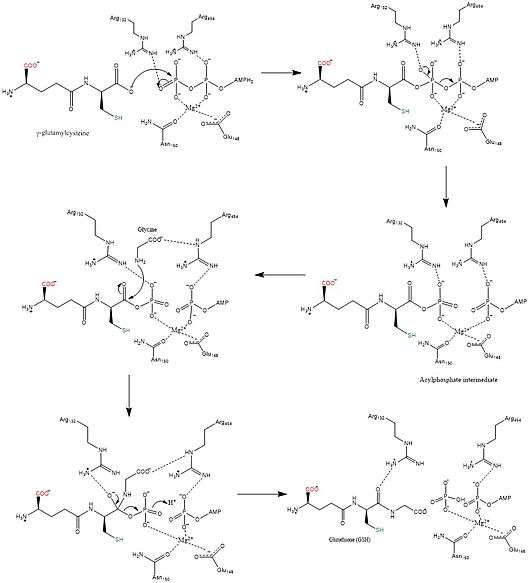
Mechanism
The biosynthetic mechanisms for synthetases use energy from nucleoside triphosphates, whereas synthases do not.[12] Glutathione synthetase stays true to this rule, in that it uses the energy generated by ATP. Initially, the carboxylate group on γ-glutamylcysteine is converted into an acyl phosphate by the transfer of an inorganic phosphate group of ATP to generate an acyl phosphate intermediate. Then the amino group of glycine participates in a nucleophilic attack, displacing the phosphate group and forming GSH.[13] After the final GSH product is made, it can be used by glutathione peroxidase to neutralize reactive oxygen species (ROS) such as H2O2 or Glutathione S-transferases in the detoxification of xenobiotics.[7]
Function
Glutathione synthetase is important for a variety of biological functions in multiple organisms. In Arabidopsis thaliana, low levels of glutathione synthetase have resulted in increased vulnerability to stressors such as heavy metals, toxic organic chemicals, and oxidative stress.[15] The presence of a thiol functional group allows its product GSH to serve both as an effective oxidizing and reducing agent in numerous biological scenarios. Thiols can easily accept a pair of electrons and become oxidized to disulfides, and the disulfides can be readily reduced to regenerate thiols. Additionally, the thiol side chain of cysteines serve as potent nucleophiles and react with oxidants and electrophilic species that would otherwise cause damage to the cell.[16] Interactions with certain metals also stabilize thiolate intermediates.[17]
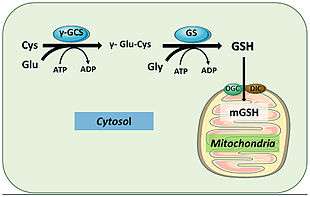
In humans, glutathione synthetase functions in a similar manner. Its product GSH participates in cellular pathways involved in homeostasis and cellular maintenance. For instance, glutathione peroxidases catalyze the oxidation of GSH to glutathione disulfide (GSSG) by reducing free radicals and reactive oxygen species such as hydrogen peroxide.[18] Glutathione S-transferase uses GSH to clean up various metabolites, xenobiotics, and electrophiles to mercapturates for excretion.[19] Because of its antioxidant role, GSS mostly produce GSH inside the cytoplasm of liver cells and imported to mitochondria where detoxification occurs.[20]]] GSH is also essential for the activation of the immune system to generate robust defense mechanisms against invading pathogens.[19] GSH is capable of preventing infection from the influenza virus.[21][22]
Clinical significance
Patients with mutations in the GSS gene develop glutathione synthetase (GSS) deficiency, an autosomal recessive disorder.[23] Patients develop a wide range of symptoms depending on the severity of the mutations. Mildly affected patients experience a compensated haemolytic anaemia because mutations affect stability of the enzyme. Moderately and severely affected individuals have enzymes with dysfunctional catalytic sites, rendering it unable to participate in detoxification reactions. Physiological symptoms include metabolic acidosis, neurological defects, and increased susceptibility to pathogenic infections.[4]
Treatment of individuals with glutathione synthetase deficiency generally involve therapeutic treatments to address mild to severe symptoms and conditions. In order to treat metabolic acidosis, severely affected patients are given large amounts of bicarbonate and antioxidants such as vitamin E and vitamin C.[24] In mild cases, ascorbate and N-acetylcysteine have been shown to increase glutathione levels and increase erythrocyte production.[25] It is important to note that because glutathione synthetase deficiency is so rare, it is poorly understood. The disease also appears on a spectrum, so it is even more difficult to generalize among the few cases that occur.[26]
See also
- Glutathione synthetase deficiency
- Glutathione
- Liver
- Sulfur Metabolism
References
- ↑ Gogos A, Shapiro L (Dec 2002). "Large conformational changes in the catalytic cycle of glutathione synthase". Structure. 10 (12): 1669–76. doi:10.1016/S0969-2126(02)00906-1. PMID 12467574.
- ↑ Njålsson R, Norgren S (2005). "Physiological and pathological aspects of GSH metabolism.". Acta Paediatr. 94 (2): 132–7. doi:10.1080/08035250410025285. PMID 15981742.
- 1 2 Li H, Xu H, Graham DE, White RH (Aug 2003). "Glutathione synthetase homologs encode alpha-L-glutamate ligases for methanogenic coenzyme F420 and tetrahydrosarcinapterin biosyntheses". Proceedings of the National Academy of Sciences of the United States of America. 100 (17): 9785–90. doi:10.1073/pnas.1733391100. PMC 187843
 . PMID 12909715.
. PMID 12909715. - 1 2 Njålsson R (Sep 2005). "Glutathione synthetase deficiency". Cellular and Molecular Life Sciences. 62 (17): 1938–45. doi:10.1007/s00018-005-5163-7. PMID 15990954.
- ↑ O'Neill M. "Glutathione Synthetase Deficiency". Online Mendelian Inheritance in Man.
- 1 2 3 Polekhina G, Board PG, Gali RR, Rossjohn J, Parker MW (Jun 1999). "Molecular basis of glutathione synthetase deficiency and a rare gene permutation event". The EMBO Journal. 18 (12): 3204–13. doi:10.1093/emboj/18.12.3204. PMC 1171401. PMID 10369661.
- 1 2 Banerjee R (2007). "Antioxidant Molecules and Redox Factors". Redox Biochemistry. Hoboken, N.J.: Wiley. p. 16. ISBN 978-0-471-78624-5.
- ↑ Fawaz MV, Topper ME, Firestine SM (Dec 2011). "The ATP-grasp enzymes". Bioorganic Chemistry. 39 (5-6): 185–91. doi:10.1016/j.bioorg.2011.08.004. PMC 3243065. PMID 21920581.
- ↑ Fyfe PK, Alphey MS, Hunter WN (Apr 2010). "Structure of Trypanosoma brucei glutathione synthetase: domain and loop alterations in the catalytic cycle of a highly conserved enzyme". Molecular and Biochemical Parasitology. 170 (2): 93–9. doi:10.1016/j.molbiopara.2009.12.011. PMC 2845819. PMID 20045436.
- ↑ Galperin MY, Koonin EV (1997). "A diverse superfamily of enzymes with ATP-dependent carboxylate-amine/thiol ligase activity". Protein Science : a Publication of the Protein Society. 6 (12): 2639–43. doi:10.1002/pro.5560061218. PMC 2143612. PMID 9416615.
- ↑ Meister A (1978). "Current Status of the γ-Glutamyl Cycle". In Wendel A, Sies H. Functions of Glutathione in Liver and Kidney. Berlin, Heidelberg: Springer Berlin Heidelberg. p. 49. ISBN 978-3-642-67132-6.
- ↑ Hara T, Kato H, Katsube Y, Oda J (Sep 1996). "A pseudo-michaelis quaternary complex in the reverse reaction of a ligase: structure of Escherichia coli B glutathione synthetase complexed with ADP, glutathione, and sulfate at 2.0 A resolution". Biochemistry. 35 (37): 11967–74. doi:10.1021/bi9605245. PMID 8810901.
- ↑ "Synthases and Ligases". IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN), and Nomenclature Commission of IUB (NC-IUB), Newsletter. 1984.
- ↑ Herrera K, Cahoon RE, Kumaran S, Jez J (Jun 2007). "Reaction mechanism of glutathione synthetase from Arabidopsis thaliana: site-directed mutagenesis of active site residues". The Journal of Biological Chemistry. 282 (23): 17157–65. doi:10.1074/jbc.M700804200. PMID 17452339.
- ↑ Moyer AM, Sun Z, Batzler AJ, Li L, Schaid DJ, Yang P, Weinshilboum RM (Mar 2010). "Glutathione pathway genetic polymorphisms and lung cancer survival after platinum-based chemotherapy". Cancer Epidemiology, Biomarkers & Prevention. 19 (3): 811–21. doi:10.1158/1055-9965.EPI-09-0871. PMC 2837367. PMID 20200426.
- ↑ Xiang C, Werner BL, Christensen EM, Oliver DJ (Jun 2001). "The biological functions of glutathione revisited in arabidopsis transgenic plants with altered glutathione levels". Plant Physiology. 126 (2): 564–74. doi:10.1104/pp.126.2.564. PMC 111149. PMID 11402187.
- ↑ Conte ML, Carroll KS (14 February 2013). "The Chemistry of Thiol Oxidation and Detection" (PDF). Oxidative Stress and Redox Regulation. pp. 1–42. doi:10.1007/978-94-007-5787-5_1.
- ↑ Suzuki N, Higuchi T, Nagano T (Aug 2002). "Multiple active intermediates in oxidation reaction catalyzed by synthetic heme-thiolate complex relevant to cytochrome p450". Journal of the American Chemical Society. 124 (32): 9622–8. doi:10.1021/ja0115013. PMID 12167058.
- 1 2 Fang YZ, Yang S, Wu G (Oct 2002). "Free radicals, antioxidants, and nutrition". Nutrition. 18 (10): 872–9. doi:10.1016/S0899-9007(02)00916-4. PMID 12361782.
- 1 2 Ribas V, García-Ruiz C, Fernández-Checa JC (Jul 2014). "Glutathione and mitochondria". Frontiers in Pharmacology. 5: 151. doi:10.3389/fphar.2014.00151. PMC 4079069. PMID 25024695.
- ↑ Townsend DM, Tew KD, Tapiero H (2003). "The importance of glutathione in human disease". Biomedicine & Pharmacotherapy = Biomédecine & Pharmacothérapie. 57 (3-4): 145–55. doi:10.1016/S0753-3322(03)00043-X. PMID 12818476.
- ↑ Cai J, Chen Y, Seth S, Furukawa S, Compans RW, Jones DP (Apr 2003). "Inhibition of influenza infection by glutathione". Free Radical Biology & Medicine. 34 (7): 928–36. doi:10.1016/S0891-5849(03)00023-6. PMID 12654482.
- ↑ Ristoff E, Mayatepek E, Larsson A (Jul 2001). "Long-term clinical outcome in patients with glutathione synthetase deficiency". The Journal of Pediatrics. 139 (1): 79–84. doi:10.1067/mpd.2001.114480. PMID 11445798.
- ↑ Kraut JA, Madias NE (May 2010). "Metabolic acidosis: pathophysiology, diagnosis and management". Nature Reviews. Nephrology. 6 (5): 274–85. doi:10.1038/nrneph.2010.33. PMID 20308999.
- ↑ Jain A, Buist NR, Kennaway NG, Powell BR, Auld PA, Mårtensson J (Feb 1994). "Effect of ascorbate or N-acetylcysteine treatment in a patient with hereditary glutathione synthetase deficiency". The Journal of Pediatrics. 124 (2): 229–33. doi:10.1016/S0022-3476(94)70309-4. PMID 8301428.
- ↑ Ristoff E, Larsson A (2007). "Inborn errors in the metabolism of glutathione". Orphanet Journal of Rare Diseases. 2: 16. doi:10.1186/1750-1172-2-16. PMC 1852094. PMID 17397529.
External links
- Glutathione Synthetase at the US National Library of Medicine Medical Subject Headings (MeSH)
