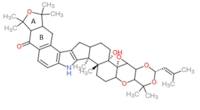
Lolitrem B
 | |
| Names | |
|---|---|
| IUPAC name
(3S,4aR,4bR,5aS,5bS,16bS,18aS,19aS)-5b-Hydroxy-1,1,10,10,12,12,16b,16c-octamethyl-3-(2-methyl-1-propen-1-yl)-1,4a,4b,6,7,7a,9,9a,10,12,12a,16,16b,16c,17,18,18a,19a-octadecahydro-5bH-[2]benzofuro[5,6-e][1,3]dioxino[5,4:2',3']oxireno[4',4a']chromeno[5',6':6,7]indeno[1,2-b]indol-13(8H)-one | |
| Identifiers | |
| 81771-19-9 | |
| 3D model (Jmol) | Interactive image |
| ChemSpider | 2342837 |
| KEGG | C20551 |
| PubChem | 11947793 |
| |
| |
| Properties | |
| C42H55NO7 | |
| Molar mass | 685.8886 |
| Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
| Infobox references | |
Lolitrem B is one of many toxins produced by a fungus called Epichloë festucae (var. lolii), which grows in perennial ryegrass. The fungus is symbiotic with the ryegrass; it doesn't harm the plant, and the toxins it produces kill insects that feed on ryegrass. Lolitrem B is one of these toxins, but it is also harmful to mammals. The shoots and flowers of infected ryegrass have especially high concentrations of lolitrem B, and when livestock eat too much of them, they get perennial ryegrass staggers. At low doses the animals have tremors, and at higher doses they stagger, and at higher yet doses the animals become paralyzed and die. The blood pressure of the animals also goes up. The effect of the lolitrem B comes on slowly and fades out slowly, as it is stored in fat after the ryegrass is eaten. The condition is especially common in New Zealand and Australia, and plant breeders there have been trying to develop strains of fungus that produce toxins only harmful to pests, and not to mammals.
Lolitrem B affects a kind of ion channel called BK channels. These channels normally open temporarily to allow neurons and other electrically-sensitive cells, like some heart cells, to "reset" after they fire; lolitrem B blocks them, preventing the neuron or heart cell from firing again. This affects nerve and heart function. The channel is also involved in blood vessel relaxation, and blocking the channel causes blood vessels to constrict, raising blood pressure.
Etymology
The Lolitrem B toxin derives the first part of its name ('Loli') from the source of the fungus (perennial ryegrass, also known as Lolium perenne), the middel part ('trem') due to the tremors the toxin is known to cause, and the last part of its name ('B') as part of a way to distinguish between different Lolitrems, based on their difference in chemical structure (see 'Chemistry').
Sources
Lolitrem B is found in perennial ryegrass that has been infected with the fungus Epichloë festucae (var. lolii).[1][2] This fungus is an endophyte; for part of its lifecycle it lives inside plants, growing between the plant cells; it is most prevalent in the ryegrass stem.[1] The fungus produces lolitrem B, one of several mycotoxins that kill pests but which also can be neurotoxins for mammals.[1]
Toxicity
When animals eat ryegrass stems infected with Epichloë festucae (var. lolii) they get a condition called perennial ryegrass staggers; in cases of mild poisoning the animals get tremors, and in severe poisoning they stagger and collapse.[1][3] In horses, tremors of the eyeball muscles are seen which are more severe during eating and exercise.[4] Lolitrem B can also increase the heart rate, blood pressure, respiration rate and disrupt the digestion process.[5][6] Lolitrems distinguish themselves among tremorgenic neurotoxins because they induce a long lasting effect on motor function and heart rate. The tremors can last for hours and at high concentrations they can cause death. In animals, lolitrem B more often causes death related to unfortunate accidents such as falling in a pond. The neurotoxic effects can be completely reversed.[7]
The threshold for toxicity varies between species of animals: for sheep a threshold value of 1.8 - 2.0 mg/kg was found, and for cattle 1.55 mg/kg.[3][7] Measuring the lolitrem B concentration in fat tissue can be used to estimate the amount of lolitrem B consumed, and is used to determine the cause of death for cattle that presenting with neurological symptoms.[6][8] Lolitrem B likely acts synergistically with ergotamine to increase smooth muscle contraction.[9]
Epidemiology
Epichloë festucae (var. lolii) infects ryegrass worldwide, but cases of perennial ryegrass staggers are rare outside of Australia and New Zealand; the reasons for this are unclear but may have to do with the purposeful selection of endophyte-infected ryegrass by plant breeders, who prize its resistance to pests which are more prevalent in Australia and New Zealand than elsewhere, and the practice of monoculture by farmers in those countries.[1]
Prevention
Plant breeders have been working with mycologists in Australia and New Zealand to develop strains of fungus that produce mycotoxins that are toxic to pests but not to mammals.[1] Until those become commercially established the best prevention is avoiding grazing livestock on ryegrass when the stems are emerging and while the plant is flowering (concentrations are highest in the mature inflorescence and in the base of the plant), and avoiding overgrazing; once the exposure to lolitrem B ends the symptoms gradually decrease.[1]
Pharmacology
Lolitrem B is rapidly eliminated from serum and has a half-life of 14 minutes.[10] Lolitrem B is not very soluble, and is generally stored in fat after ingestion and slowly released; this is likely why its effects come on slowly and linger after ingestion has stopped.[1] The more that is ingested, the more is stored in fat.[11][12]
Lolitrem B targets the large conductance calcium-activated potassium channels (BK channels) and in particular the α subunit (hSlo) of the BK channels.[1] These channels open temporarily to allow neurons to "reset" after they fire; lolitrem B blocks them, preventing the neuron from firing again after it depolarizes, which at low doses leads to tremors and at high doses to paralysis and death.[1]
The binding site of lolitrem B is likely to be located in this α subunit.[13][14] When lolitrem B is added, the potassium current quickly gets abolished and this inhibition cannot be reversed by washout (this reversal is possible for paxilline). However, over time lolitrem B slowly dissociates from the binding site. The inhibition by lolitrem B is calcium concentration-dependent. The concentration with half of the maximal inhibition (IC50) for hSlo was found to be 3.7 ± 0.4 nM. Lolitrem B is a more potent neurotoxin in vitro compared to paxilline.[10][13][14]
Lolitrem B preferably blocks the open configuration of BK channels, as under high calcium concentrations promoting the opening of BK channels, the apparent affinity increases three-fold.[15][16] The inhibition by lolitrem B and its affinity differs with the calcium concentration. Lolitrem B has the highest affinity for BK channels when there is a high probability of an open conformation thus when the calcium binds to the high affinity sites. The inhibition occurs when the channels are in an open state.[15]
BK channels oppose vasoconstriction in blood vessels resulting in vasorelaxation. Blocking the channels leads to vasoconstriction and to an increase in blood pressure.[17] The BK channel α subunit is expressed in muscle and nerve tissue and the BK channels are abundant in the brain.[18] The BK channels modulate neurotransmitter release, the form of the action potential and repetitive firing.[19] Inhibition of the channels can explain why there would be an increased release in excitatory neurotransmitters resulting in tremors, ataxia, hypersensitivity, increased smooth muscle contraction in the colon and an increased heart rate.[5][9][20]
Chemistry
Lolitrem B is the most potent member of the lolitrem family.[1] It possesses an indole-diterpene unit as well as a reactive epoxide group.[21]
It structurally looks like paxilline which is a related tremor inducer.[22] There are multiple lolitrems which are labelled by a letter. The difference between them is the position and number of aryl and hydroxyl substituents plus the absence or presence of an I ring. The I ring seems to be necessary for prolonged tremors to occur.[23] Intermediate metabolites such as terpendoles and paspaline can become lolitrems by addition of two rings (A and B) at the C20-C21 position to the indole moiety of the molecule.[24]
Biosynthesis
The production of lolitrem B requires 10 different genes on a locus (the LTM locus) which is organized in three clusters. These clusters are separated by large AT-rich sequences. Cluster 1 contains the genes ltmG, ltmK and ltmM. Cluster 2 contains ltmP, ltmF, ltmB, ltmQ and ltmC and cluster 3 ltmE and ltmJ Four genes from cluster 2 are orthologues of functional characterized paxilline genes, meaning that the genes show homologous sequences. The genes in cluster 3 appear to be unique to the Epichloë genus.[25][26]
See also
- Penitrem A - a structurally related fungal neurotoxin found on ryegrass
References
- 1 2 3 4 5 6 7 8 9 10 11 Philippe, G (15 February 2016). "Lolitrem B and Indole Diterpene Alkaloids Produced by Endophytic Fungi of the Genus Epichloë and Their Toxic Effects in Livestock.". Toxins. 8 (2): 47. PMC 4773800
 . PMID 26891327.
. PMID 26891327. - ↑ Note: This fungus was formerly called Neotyphodium lolii. See Leuchtmann, Adrian; Bacon, Charles W.; Schardl, Christopher L.; White, James F.; Tadych, Mariusz (2014-03-01). "Nomenclatural realignment of Neotyphodium species with genus Epichloë". Mycologia. 106 (2): 202–215. doi:10.3852/13-251. PMID 24459125.
- 1 2 Cheeke, P R (2016-10-11). "Endogenous toxins and mycotoxins in forage grasses and their effects on livestock.". Journal of Animal Science. 73 (3). doi:10.2527/1995.733909x. ISSN 0021-8812.
- ↑ Johnstone, L. K.; Mayhew, I. G.; Fletcher, L. R. (2012-05-01). "Clinical expression of lolitrem B (perennial ryegrass) intoxication in horses". Equine Veterinary Journal. 44 (3): 304–309. doi:10.1111/j.2042-3306.2011.00439.x. ISSN 2042-3306.
- 1 2 McLEAY, L. M; SMITH, B. L; MUNDAY-FINCH, S. C (1999-04-01). "Tremorgenic mycotoxins paxilline, penitrem and lolitrem B, the non-tremorgenic 31-epilolitrem B and electromyographic activity of the reticulum and rumen of sheep". Research in Veterinary Science. 66 (2): 119–127. doi:10.1053/rvsc.1998.0253.
- 1 2 Fisher, M. J.; Bohnert, D. W.; Ackerman, C. J.; Schauer, C. S.; DelCurto, T.; Craig, A. M.; Vanzant, E. S.; Harmon, D. L.; Schrick, F. N. (2004-07-01). "Evaluation of perennial ryegrass straw as a forage source for ruminants". Journal of Animal Science. 82 (7). doi:10.2527/2004.8272175x. ISSN 1525-3163.
- 1 2 Gallagher, R. T.; Hawkes, A. D. "The potent tremorgenic neurotoxins lolitrem B and aflatrem: A comparison of the tremor response in mice". Experientia. 42 (7): 823–825. doi:10.1007/BF01941539. ISSN 0014-4754.
- ↑ Tor-Agbidye, J.; Blythe, L. L.; Craig, A. M. (2001-06-01). "Correlation of endophyte toxins (ergovaline and lolitrem B) with clinical disease: fescue foot and perennial ryegrass staggers". Veterinary and Human Toxicology. 43 (3): 140–146. ISSN 0145-6296. PMID 11383653.
- 1 2 Dalziel, J. E.; Dunstan, K. E.; Finch, S. C. (2013-11-01). "Combined effects of fungal alkaloids on intestinal motility in an in vitro rat model". Journal of Animal Science. 91 (11): 5177–5182. doi:10.2527/jas.2013-6449. ISSN 1525-3163.
- 1 2 "Distribution Du Lolitrème B et de L'ergovaline Après Un Bolus Intraruminal Chez La Chèvre En Lactation" (PDF). Rencontres Recherches Ruminants. 11.
- ↑ Miyazaki, Shigeru; Ishizaki, Ikumi; Ishizaka, Masumi; Kanbara, Tadashi; Ishiguro-Takeda, Yuriko (2004-07-01). "Lolitrem B Residue in Fat Tissues of Cattle Consuming Endophyte-Infected Perennial Ryegrass Straw". Journal of Veterinary Diagnostic Investigation. 16 (4): 340–342. doi:10.1177/104063870401600416. ISSN 1040-6387.
- ↑ Finch, S. C.; Fletcher, L. R.; Babu, J. V. (2012-01-01). "The evaluation of endophyte toxin residues in sheep fat". New Zealand Veterinary Journal. 60 (1): 56–60. doi:10.1080/00480169.2011.634746. ISSN 0048-0169.
- 1 2 Dalziel, Julie E.; Finch, Sarah C.; Dunlop, James (2005-03-15). "The fungal neurotoxin lolitrem B inhibits the function of human large conductance calcium-activated potassium channels". Toxicology Letters. 155 (3): 421–426. doi:10.1016/j.toxlet.2004.11.011.
- 1 2 Imlach, Wendy L.; Finch, Sarah C.; Dunlop, James; Meredith, Andrea L.; Aldrich, Richard W.; Dalziel, Julie E. (2008-11-12). "The Molecular Mechanism of "Ryegrass Staggers," a Neurological Disorder of K Channels". Journal of Pharmacology and Experimental Therapeutics. 327 (3): 657–664. doi:10.1124/jpet.108.143933.
- 1 2 Imlach, Wendy L.; Finch, Sarah C.; Zhang, Yanli; Dunlop, James; Dalziel, Julie E. (2011-04-01). "Mechanism of action of lolitrem B, a fungal endophyte derived toxin that inhibits BK large conductance Ca2+-activated K+ channels". Toxicon. 57 (5): 686–694. doi:10.1016/j.toxicon.2011.01.013.
- ↑ Zeng, Xu-Hui; Xia, Xiao-Ming; Lingle, Christopher J. (2005-03-01). "Divalent Cation Sensitivity of BK Channel Activation Supports the Existence of Three Distinct Binding Sites". The Journal of General Physiology. 125 (3): 273–286. doi:10.1085/jgp.200409239. ISSN 0022-1295.
- ↑ Brayden, J. E.; Nelson, M. T. (1992-04-24). "Regulation of arterial tone by activation of calcium-dependent potassium channels". Science. 256 (5056): 532–535. doi:10.1126/science.1373909. ISSN 0036-8075.
- ↑ Knaus, H.G. "Distribution of high-conductance Ca2+-activated K+ channels in rat brain: targeting to axons and nerve terminals". The Journal.of Neuroscience. 16.
- ↑ Shao, Li-Rong; Halvorsrud, Ragnhild; Borg-Graham, Lyle; Storm, Johan F. (1999-11-01). "The role of BK-type Ca2+-dependent K+ channels in spike broadening during repetitive firing in rat hippocampal pyramidal cells". The Journal of Physiology. 521 (1): 135–146. doi:10.1111/j.1469-7793.1999.00135.x. ISSN 1469-7793.
- ↑ Mantle, P.G. "Amino acid neurotransmitter release from cerebrocortical synaptosomes of sheep with severe ryegrass staggers in New Zealand". Research in Veterinary Science. 34.
- ↑ Gallagher, R. T.; White, E. P.; Mortimer, P. H. (1981-10-01). "Ryegrass Staggers: Isolation of Potent Neurotoxins Lolitrem a and Lolitrem B From Staggers-Producing Pastures". New Zealand Veterinary Journal. 29 (10): 189–190. doi:10.1080/00480169.1981.34843. ISSN 0048-0169.
- ↑ Miles, Christopher O.; Wilkins, Alistair L.; Gallagher, Rex T.; Hawkes, Allan D.; Munday, Sarah C.; Towers, Neale R. (1992-02-01). "Synthesis and tremorgenicity of paxitriols and lolitriol: possible biosynthetic precursors of lolitrem B". Journal of Agricultural and Food Chemistry. 40 (2): 234–238. doi:10.1021/jf00014a013. ISSN 0021-8561.
- ↑ Munday-Finch, Sarah C.; Wilkins, Alistair L.; Miles, Christopher O.; Tomoda, Hiroshi; Ōmura, Satoshi (1997-01-01). "Isolation and Structure Elucidation of Lolilline, a Possible Biosynthetic Precursor of the Lolitrem Family of Tremorgenic Mycotoxins". Journal of Agricultural and Food Chemistry. 45 (1): 199–204. doi:10.1021/jf960396r. ISSN 0021-8561.
- ↑ Young, Carolyn A.; Tapper, Brian A.; May, Kimberley; Moon, Christina D.; Schardl, Christopher L.; Scott, Barry (2009-04-01). "Indole-Diterpene Biosynthetic Capability of Epichloë Endophytes as Predicted by ltm Gene Analysis". Applied and Environmental Microbiology. 75 (7): 2200–2211. doi:10.1128/AEM.00953-08. ISSN 0099-2240.
- ↑ Young, Carolyn A.; Felitti, Silvina; Shields, Katherine; Spangenberg, German; Johnson, Richard D.; Bryan, Gregory T.; Saikia, Sanjay; Scott, Barry (2006-10-01). "A complex gene cluster for indole-diterpene biosynthesis in the grass endophyte Neotyphodium lolii". Fungal Genetics and Biology. 43 (10): 679–693. doi:10.1016/j.fgb.2006.04.004.
- ↑ Young, C. A.; Bryant, M. K.; Christensen, M. J.; Tapper, B. A.; Bryan, G. T.; Scott, B. (2005-07-01). "Molecular cloning and genetic analysis of a symbiosis-expressed gene cluster for lolitrem biosynthesis from a mutualistic endophyte of perennial ryegrass". Molecular Genetics and Genomics. 274 (1): 13–29. doi:10.1007/s00438-005-1130-0. ISSN 1617-4615.
| Bacterial toxins |
| ||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mycotoxins |
| ||||||||||||||||||||||||||||||||||||||||||
| Plant toxins |
| ||||||||||||||||||||||||||||||||||||||||||
| Invertebrate toxins | |||||||||||||||||||||||||||||||||||||||||||
| Vertebrate toxins |
| ||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||